Azione complementare dei geni. Interazione di geni non allelici: complementarità, epistasi, polimerizzazione, pleiotropia Interazione genica di complementarità
Leggi anche
Interazione dei geni allelici e suoi tipi
Studiando le leggi di G. Mendel, abbiamo già conosciuto i principali tipi di interazione dei geni allelici. Sulla base del materiale precedentemente studiato, compila la tabella.
Tavolo. Tipi di interazione tra geni di una coppia allelica
Tipo di interazione |
La natura dell'interazione in |
Formule per la scissione fenotipica in F 2 e negli incroci test |
Esempi |
| Dominio completo | Un allele (dominante) sopprime completamente l'effetto di un altro allele (recessivo). |
L'alta statura delle piante domina il nanismo; capelli ricci - sopra i capelli lisci; occhi marroni - sopra quelli blu, ecc. |
|
| Dominanza incompleta | L'effetto di ciascun allele non è pienamente realizzato, a seguito del quale appare un tratto intermedio |
Colorazione dei fiori della bellezza notturna; anemia falciforme; cistonuria, ecc. |
|
| Codominanza | Partecipazione di entrambi gli alleli nella determinazione del tratto |
Gruppo sanguigno IV (gli eritrociti di questo gruppo sanguigno hanno agglutinogeni del tipo UN e tipo IN) |
Interazione di geni non allelici
Sulla base delle leggi di G. Mendel, possiamo giungere alla conclusione che esiste una connessione abbastanza forte tra un gene e un tratto, che il genotipo è costituito dalla somma di geni che agiscono indipendentemente e il fenotipo è una combinazione meccanica dei tratti individuali. Tuttavia, connessioni dirette e inequivocabili tra un gene e un tratto sono l’eccezione piuttosto che la regola. Lo sviluppo di un tratto in un organismo è solitamente sotto il controllo di molti geni e il tratto è il risultato dell'interazione di geni non allelici.
Il caso più semplice di interazione di geni non allelici è quando un tratto è controllato da due coppie di alleli. Diamo un'occhiata ad alcuni esempi di tale interazione.
Interazione tra geni complementari
Il fenomeno dell'interazione genica, in cui la presenza di due geni non allelici è necessaria per la manifestazione di un tratto, è chiamato complementarità(dal lat. complemento- aggiunta) e i geni necessari per la manifestazione del tratto lo sono complementare o aggiuntivi.
Consideriamo varie opzioni per l'interazione tra geni complementari.
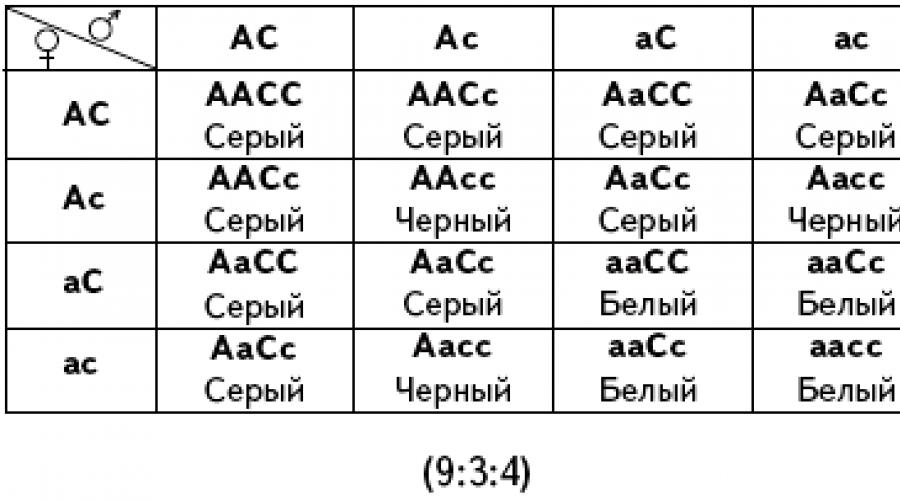
Gli alleli del secondo gene compaiono solo in presenza di un allele dominante del geneUN. La colorazione dei topi dipende, nel caso più semplice, da due geni. Topi con genotipo ahh mancano di pigmento e sono di colore bianco. In presenza di un allele dominante UN il pigmento viene prodotto e il topo viene in qualche modo colorato. Il colore specifico è determinato dal secondo gene. Il suo allele dominante CON provoca il colore grigio del topo e l'allele recessivo Con- colore nero. Pertanto, se consideriamo le varianti omozigoti, il genotipo dei topi grigi lo è AASS, nero - AAAss, bianco - aaSS O culo. Quando si incrocia un topo grigio AASS con il bianco culo nella prima generazione tutto risulta secondo Mendel: tutti gli ibridi sono di colore grigio (questi sono eterozigoti con il genotipo AaSs). In F2, come è facile verificare, otteniamo 9/16 topi grigi, 3/16 neri e 4/16 bianchi.

Perché un tratto si manifesti, nel genotipo devono essere presenti alleli dominanti di due geni diversi. Un esempio di tale interazione genetica complementare è l'ereditarietà della forma del frutto nelle zucche. In presenza di entrambi gli alleli dominanti, i frutti hanno una forma a forma di disco, in presenza di un (qualsiasi!) allele dominante - sferico e in assenza di alleli dominanti - allungato.

Perché un tratto si manifesti, devono essere presenti nel genotipo gli alleli dominanti di due geni diversi, ma ciascun allele dominante, in combinazione con gli alleli recessivi dell'altra coppia, ha una manifestazione fenotipica indipendente. Ad esempio, nei polli, la cresta a forma di pisello è determinata da un gene dominante, mentre la cresta a forma di rosa è determinata da un altro gene non allelico, ma anch'esso dominante. Quando questi geni appartengono allo stesso genotipo, si sviluppa una cresta a forma di nocciola. Se l'organismo è omozigote per entrambi i geni recessivi, si sviluppa un semplice pettine a forma di foglia. Quando si incrociano i diibridi (tutti con cresta a forma di noce), nella seconda generazione avviene una scissione in un rapporto di 9: 3: 3: 1. Ma qui è impossibile trovare una segregazione indipendente di ciascun allele in un rapporto 3:1, poiché in tutti i casi di coincidenza nel genotipo di entrambi i geni dominanti, il loro effetto diretto non viene rilevato.
Pertanto, parliamo di interazione complementare di geni non allelici nel caso in cui due geni non allelici forniscano un nuovo tratto, cioè si verifichi una nuova formazione.
Interazione del gene epistatico
Viene chiamata l'interazione dei geni, in cui un gene sopprime l'azione di un altro, non allelico al primo epistasi(dal greco epi– sopra e stasi- stare in piedi) e vengono chiamati i geni che sopprimono l'azione di altri geni geni soppressori, O geni inibitori(dal lat. inibitore– tenere premuto), oppure geni epistatici. Il gene che viene soppresso si chiama ipostatico.
Esistono epistasi dominanti e recessive.
Epistasi dominante associato ad un gene inibitore dominante. Ad esempio, gene IO determina il colore bianco dei frutti di zucca; se presente, l'effetto del gene IN non appare. Con genotipo iiBB O iiВb I frutti della zucca sono gialli. Infine, se entrambi i geni sono rappresentati da alleli recessivi, il frutto della zucca è verde. Lo stesso tipo può essere utilizzato per ereditare il colore del mantello di cani, pecore e altri animali.
Epistasi recessiva viene rilevato quando l'espressione di un gene viene soppressa da alleli recessivi di un altro gene. Questo tipo di epistasi è illustrato dall'ereditarietà del colore del mantello nei topi domestici. Il colore Agouti (grigio-rossastro) è determinato dal gene dominante UN, il suo allele recessivo UN dà un colore nero. Gene da una coppia non allelica IN promuove la manifestazione del colore e del gene Bè un soppressore e sopprime l'effetto dell'allele dominante UN e recessivo UN. Topi con genotipo Abb indistinguibili nel fenotipo dagli individui con il genotipo aabb- tutto bianco.
Interazione tra geni polimerici
Polimerismo– un tipo di interazione di geni non allelici in cui diverse coppie di geni non allelici agiscono sullo sviluppo di un tratto. Tali geni sono chiamati polimero(dal greco politica– molto e meros- Parte). Sono indicati con la stessa lettera, ma con indici diversi, che indicano il numero di coppie alleliche nel genotipo che determinano lo sviluppo di un particolare tratto ( A1A1A2A2).
Esistono due varianti di polimerizzazione: con l'effetto somma dei geni e senza il potenziamento dei geni tra loro.
L'effetto cumulativo dei geni polimerici. In questo caso, il grado di manifestazione del tratto dipende dal numero di alleli dominanti nel genotipo dell’individuo. Pertanto, il colore rosso dei chicchi di grano è determinato da due o più coppie di geni. Ciascuno dei geni dominanti di questi alleli determina il colore rosso, mentre i geni recessivi determinano il colore bianco dei chicchi. Un allele dominante produce grani non molto colorati. Se nel genotipo sono presenti due alleli dominanti, l'intensità del colore aumenta. Solo quando l'organismo è omozigote per tutte le coppie di geni recessivi i grani non sono colorati. Pertanto, quando si incrociano i diibridi, si verifica una divisione nel rapporto tra 15 colorati e uno bianco. Ma su 15 dipinti, uno avrà un colore rosso intenso, perché... contiene quattro alleli dominanti, quattro saranno di colore leggermente più chiaro, perché contengono tre alleli dominanti e un allele recessivo nei genotipi, sei sono ancora più leggeri con due alleli dominanti e due recessivi nei genotipi, quattro sono ancora più leggeri, perché hanno solo un gene dominante e tre recessivi, cioè la vera divisione sarà 1: 4: 6: 4: 1.
Secondo questa versione del polimero, negli esseri umani il colore della pelle, l'altezza e il peso vengono ereditati. Un simile meccanismo di ereditarietà è caratteristico di molti tratti quantitativi, compresi quelli economicamente preziosi: contenuto di zucchero nelle radici di barbabietola, contenuto vitaminico in frutta e verdura, lunghezza della spiga di cereali, lunghezza della spiga di mais, fertilità animale, produzione di latte nel bestiame , produzione di uova di polli, ecc.
I geni dei polimeri non si potenziano a vicenda. In questo caso la suddivisione in Fa 2 sarà 15:1. Pertanto, i frutti della borsa del pastore possono essere triangolari (dominanti) e ovali (recessivi). Il tratto è controllato da due coppie di geni polimerici. Se il genotipo di una pianta ha almeno un allele dominante della prima o della seconda coppia di geni polimerici, la forma del frutto sarà triangolare ( A1A2; A1 e 2; un 1 A 2 ). Solo le piante che non hanno un solo allele dominante nel loro genotipo avranno un frutto di forma ovale - un 1 un 1 un 2 un 2.
Pertanto, l'accumulo di determinati alleli nel genotipo può portare a cambiamenti nell'espressione dei tratti.
Azione genetica multipla
 Spesso un gene influenza non uno, ma una serie di tratti di un organismo. Viene chiamato il fenomeno in cui un gene può influenzare la formazione di diverse caratteristiche di un organismo pleiotropia(dal greco pleone– più numerosi e tropos- giro).
Spesso un gene influenza non uno, ma una serie di tratti di un organismo. Viene chiamato il fenomeno in cui un gene può influenzare la formazione di diverse caratteristiche di un organismo pleiotropia(dal greco pleone– più numerosi e tropos- giro).
L’esistenza di questo fenomeno non contraddice affatto il concetto classico di “un gene – una proteina – un tratto”. In modo semplificato, l'influenza di un gene su più tratti contemporaneamente può essere rappresentata come segue. Come risultato della lettura delle informazioni da un gene, si forma una determinata proteina, che può quindi partecipare a vari processi che si verificano nel corpo, esercitando così molteplici effetti. Ad esempio, nei piselli il colore marrone del tegumento, lo sviluppo del pigmento in altre parti della pianta e il colore dei fiori dipendono da un gene; nella Drosophila il gene che causa il colore bianco degli occhi influisce sullo schiarimento del colore del corpo e sui cambiamenti di alcuni organi interni; Nell’uomo il gene responsabile dei capelli rossi determina anche il colore più chiaro della pelle e la comparsa delle lentiggini. La sindrome di Marfan è causata da un gene pleiotropico autosomico dominante e si manifesta con crescita elevata, allungamento delle ossa delle dita delle mani e dei piedi (dita a ragno), ipermobilità articolare, sublussazione del cristallino e malattie cardiache.
Questa azione multipla è tipica della maggior parte dei geni. Tuttavia, non si deve immaginare che un gene pleiotropico influenzi equamente ogni tratto. La stragrande maggioranza dei geni con vari gradi di pleiotropia sono caratterizzati da un effetto più forte su un tratto e da un effetto molto più debole su un altro.
Genotipo come sistema integrale
Il fatto della divisione degli ibridi nella prole ci consente di affermare che il genotipo è composto da elementi individuali - geni che possono essere ereditati indipendentemente ( discrezionalità del genotipo). Allo stesso tempo, il genotipo non può essere considerato come una semplice somma meccanica di geni individuali. Un genotipo è un sistema di geni interagenti. Più precisamente, non sono i geni stessi (sezioni di DNA) che interagiscono, ma i prodotti genici formati sulla loro base (RNA e quindi proteine). Pertanto, in alcuni casi, l'azione di geni diversi è relativamente indipendente, ma, di regola, la manifestazione di un tratto è il risultato dell'interazione di prodotti di geni diversi.
Il genotipo di qualsiasi organismo è un sistema integrale complesso di geni interagenti. Questa integrità del genotipo è nata storicamente nel processo di evoluzione della specie. Come risultato delle mutazioni, compaiono costantemente nuovi geni, si formano nuovi cromosomi e persino nuovi genomi. I geni appena emersi possono interagire immediatamente con i geni esistenti o modificare la natura del lavoro di questi ultimi, anche se sono recessivi, cioè senza manifestarsi.
Di conseguenza, in ogni specie di piante e animali il genotipo si manifesta come un sistema integrale che si è sviluppato storicamente in un dato momento.
II. Consolidamento della conoscenza
Riassumere la conversazione mentre si apprende nuovo materiale.
III. Compiti a casa
Studia il paragrafo del libro di testo (relazione tra geni e tratti, tipi di interazione di geni allelici e non allelici, pleiotropia, genotipo come sistema integrale).
Risolvi i problemi alla fine del paragrafo del libro di testo.
Complementare è un tipo di interazione di geni non allelici in cui l'azione di un gene di una coppia allelica è completata dall'azione di un gene di un'altra coppia allelica, determinando la formazione di un tratto qualitativamente nuovo.
Un classico esempio di questa interazione è l'ereditarietà della forma del pettine nei polli. Si trovano le seguenti forme di pettine: a forma di foglia - il risultato dell'interazione di due geni recessivi non allelici UNabb; a forma di noce - il risultato dell'interazione di due geni non allelici dominanti UN- B-; rosa e a forma di pisello – con genotipi UN- bb E aaB- , rispettivamente.
Un altro esempio è l’ereditarietà del colore del mantello nei topi. Il colore è grigio, bianco e nero e c'è un solo pigmento: il nero. La formazione di un particolare colore del mantello si basa sull'interazione di due coppie di geni non allelici:
UN – un gene che determina la sintesi del pigmento;
UN – un gene che non determina la sintesi del pigmento;
B – un gene che determina la distribuzione non uniforme del pigmento;
B – un gene che determina la distribuzione uniforme del pigmento.
Esempi di interazione complementare nell'uomo: retinoblastoma e nefroblastoma sono codificati da due coppie di geni non allelici.
Possibili opzioni per la scissione in F 2 durante l'interazione complementare: 9:3:4; 9:3:3:1; 9:7.
Epistasi
L'epistasi è un tipo di interazione di geni non allelici in cui l'azione di un gene di una coppia allelica viene soppressa dall'azione di un gene di un'altra coppia allelica.
Esistono due forme di epistasi: dominante e recessivo. Nell'epistasi dominante, il gene dominante agisce come un gene soppressore; nell'epistasi recessiva, il gene recessivo agisce come un gene soppressore.
Un esempio di epistasi dominante è l'eredità del colore del piumaggio nei polli. Due coppie di geni non allelici interagiscono:
CON– un gene che determina il colore del piumaggio (solitamente variegato),
Con– un gene che non determina il colore del piumaggio,
IO – gene che sopprime la colorazione,
io– un gene che non sopprime il colore.
Opzioni di divisione in F 2: 12:3:1, 13:3.
Negli esseri umani, un esempio di epistasi dominante è fermentopatia (enzimopatie) sono malattie basate sulla produzione insufficiente dell'uno o dell'altro enzima.
Un esempio di epistasi recessiva è il cosiddetto “fenomeno Bombay”: in una famiglia di genitori in cui la madre aveva il gruppo sanguigno O e il padre aveva il gruppo sanguigno A, nacquero due figlie femmine, una delle quali aveva il gruppo sanguigno AB. Gli scienziati hanno suggerito che la madre avesse il gene I B nel suo genotipo, ma il suo effetto era soppresso da due geni dd epistatici recessivi.
Polimerismo
Il polimerismo è un tipo di interazione di geni non allelici in cui diversi geni non allelici determinano lo stesso tratto, migliorandone la manifestazione. Questo fenomeno è l'opposto della pleiotropia. I tratti quantitativi vengono solitamente ereditati in base al tipo di polimero, il che spiega l'ampia varietà delle loro manifestazioni in natura.
Ad esempio, il colore dei chicchi nel grano è determinato da due coppie di geni non allelici:
UN 1
UN 1 – un gene che non determina la colorazione rossa;
UN 2 – gene che determina la colorazione rossa;
UN 2 - un gene che non determina la colorazione rossa.
UN 1 UN 1 UN 2 UN 2 – genotipo di piante con chicchi di colore rosso;
UN 1 UN 1 UN 2 UN 2 - genotipo vegetale con colore bianco dei grani.
Divisione in Fa 2: 15:1 o 1:4:6:4:1.
Negli esseri umani, il tipo di polimero eredita caratteristiche quali altezza, colore dei capelli, colore della pelle, pressione sanguigna e capacità mentali.
I geni non allelici possono anche interagire tra loro. Inoltre, il loro principio di interazione è leggermente diverso dalla relazione dominante-recessiva come nel caso dei geni allelici.
È più corretto parlare non dell'interazione dei geni, ma dell'interazione dei loro prodotti, cioè dell'interazione delle proteine sintetizzate sulla base dei geni.
Interazione complementare di geni non allelici- questa è la loro interazione in cui i loro prodotti completano le azioni degli altri.
Un esempio di interazione genetica complementare è il colore degli occhi della mosca Drosophila. Le mosche con il genotipo S-B hanno normali occhi rossi, ssbb - bianchi, S-bb - marroni, ssB- - scarlatti brillanti. Pertanto, se entrambi i geni non allelici sono recessivi, non viene sintetizzato alcun pigmento e gli occhi diventano bianchi. Se è presente solo il gene dominante S, appare un pigmento marrone e solo il gene dominante B appare scarlatto brillante. Se ci sono due geni dominanti, i loro prodotti interagiscono tra loro formando il colore rosso.
Con l'interazione complementare dei geni durante l'incrocio di eterozigoti (AaBb), sono possibili diverse suddivisioni fenotipiche (9:6:1, 9:3:3:1, 9:3:4, 9:7).
Epistasi- questa è l'interazione di geni non allelici quando l'azione di un gene sopprime l'azione di un altro. Sia un allele dominante che quello recessivo di un dato gene possono avere un effetto epistatico (soppressivo) su un altro gene. La scissione del fenotipo nell'epistasi dominante differisce dall'epistasi recessiva. Un gene epistatico è solitamente indicato con la lettera I.
Un esempio di epistasi è la comparsa di piumaggio colorato nella seconda generazione quando si incrociano polli bianchi di razze diverse. Alcuni hanno il genotipo IIAA, altri hanno il genotipo iiaa. F1 - IaAa. In F 2 avviene la consueta suddivisione del genotipo: 9I-A-: 3I-aa: 3iiA-: 1iicc. In questo caso, gli uccelli con il genotipo iiA- sono colorati, il che determina il gene dominante A, che in un genitore era soppresso dal gene inibitore dominante I, e nell'altro era presente solo in forma recessiva.
A Interazione polimerica di geni non allelici il grado di espressione del tratto (la sua quantità) dipende dal numero di geni allelici e non allelici dominanti. Quanti più geni sono coinvolti nell'interazione del polimero, tanto più diversi sono i gradi di espressione del tratto. Ciò avviene durante la polimerizzazione cumulativa, quando tutti i geni partecipano all'accumulo di un tratto. Con la polimerizzazione non cumulativa, il numero di geni dominanti non influenza il grado di espressione del tratto; almeno uno è sufficiente; e una forma fenotipicamente distinta si osserva solo negli individui in cui tutti i geni polimerici sono recessivi.
Il polimerismo, ad esempio, determina il colore della pelle umana. Quattro geni (o quattro coppie di alleli secondo altre fonti) hanno un effetto. Consideriamo una situazione con due coppie. Quindi A 1 A 1 A 2 A 2 determinerà il colore più scuro e a 1 a 1 a 2 a 2 determinerà il più chiaro. Il colore medio della pelle apparirà se due geni qualsiasi sono dominanti (A 1 a 1 A 2 a 2, A 1 A 1 a 2 a 2, a 1 a 1 A 2 A 2). La presenza di un gene dominante risulterà in un colore della pelle vicino al chiaro, ma più scuro, e tre geni dominanti - vicino allo scuro, ma più chiaro.
Succede che un gene determina diversi tratti. Questa azione genetica si chiama pleiotropia. È chiaro che qui non stiamo parlando dell'interazione dei geni, ma delle molteplici azioni di un gene.
La trasmissione dei tratti di generazione in generazione è dovuta all'interazione di vari geni. Cos'è un gene e quali tipi di interazioni ci sono tra loro?
Cos'è un gene?
Attualmente per genoma si intende l'unità di trasmissione dell'informazione ereditaria. I geni si trovano nel DNA e formano le sue sezioni strutturali. Ogni gene è responsabile della sintesi di una specifica molecola proteica, che determina la manifestazione di un particolare tratto nell'uomo.
Ogni gene ha diversi sottotipi o alleli, che causano una varietà di tratti (ad esempio, il colore degli occhi marroni è causato dall'allele dominante del gene, mentre il colore blu è un tratto recessivo). Gli alleli si trovano in aree identiche e la trasmissione dell'uno o dell'altro cromosoma determina la manifestazione dell'uno o dell'altro tratto.
Tutti i geni interagiscono tra loro. Esistono diversi tipi della loro interazione: allelica e non allelica. Di conseguenza, si distingue l'interazione dei geni allelici e non allelici. In cosa differiscono l'uno dall'altro e come si manifestano?
Storia della scoperta
Prima che venissero scoperti i tipi di interazione dei geni non allelici, era generalmente accettato che fosse solo possibile (se c'è un gene dominante, apparirà il tratto; se non c'è, il tratto non esisterà). Prevalse la dottrina dell'interazione allelica, che per lungo tempo fu il dogma principale della genetica. La dominanza è stata studiata a fondo e sono stati scoperti tipi come la dominanza completa e incompleta, la co-dominanza e la sovra-dominanza.

Tutti questi principi erano soggetti al primo, che stabiliva l'uniformità degli ibridi di prima generazione.
Dopo ulteriori osservazioni e ricerche, si è notato che non tutti i segni si adattano alla teoria della dominanza. Dopo uno studio più approfondito, è stato dimostrato che non solo i geni identici influenzano la manifestazione di una caratteristica o di un gruppo di proprietà. Pertanto, sono state scoperte forme di interazione tra geni non allelici.
Reazioni tra geni
Come è stato detto, la dottrina dell’eredità dominante prevalse per molto tempo. In questo caso si è verificata un'interazione allelica in cui il tratto si manifestava solo nello stato eterozigote. Dopo che furono scoperte varie forme di interazione tra geni non allelici, gli scienziati furono in grado di spiegare tipi di eredità fino ad allora inspiegabili e ottenere risposte a molte domande.

Si è scoperto che la regolazione genetica dipende direttamente dagli enzimi. Questi enzimi hanno permesso ai geni di reagire in modi diversi. In questo caso, l'interazione dei geni allelici e non allelici è avvenuta secondo gli stessi principi e schemi. Ciò ha portato alla conclusione che l'ereditarietà non dipende dalle condizioni in cui i geni interagiscono e che la ragione della trasmissione atipica dei tratti risiede nei geni stessi.
L'interazione non allelica è unica, il che consente di ottenere nuove combinazioni di caratteristiche che determinano un nuovo grado di sopravvivenza e sviluppo degli organismi.
Geni non allelici

I geni non allelici sono quelli localizzati in diverse regioni di cromosomi non omologhi. Hanno la stessa funzione di sintesi, ma codificano per la formazione di proteine diverse che determinano caratteristiche diverse. Tali geni, reagendo tra loro, possono determinare lo sviluppo di tratti in diverse combinazioni:
- Un tratto sarà determinato dall'interazione di diversi geni che hanno una struttura completamente diversa.
- Diversi tratti dipenderanno da un singolo gene.
Le reazioni tra questi geni sono un po' più complesse rispetto all'interazione allelica. Tuttavia, ciascuno di questi tipi di reazioni ha le sue caratteristiche e caratteristiche.
Quali tipi di interazione dei geni non allelici esistono?
- Epistasi.
- Polimerismo.
- Complementarità.
- Azione dei geni modificatori.
- Interazione pleiotropica.
Ciascuno di questi tipi di interazione ha le sue proprietà uniche e si manifesta a modo suo.
Vale la pena soffermarsi più in dettaglio su ciascuno di essi.
Epistasi
Questa interazione di geni non allelici - epistasi - si osserva quando un gene sopprime l'attività di un altro (il gene soppressore è chiamato gene epistatico e il gene soppresso è chiamato gene ipostatico).
La reazione tra questi geni può essere dominante o recessiva. L'epistasi dominante si osserva quando un gene epistatico (solitamente indicato con la lettera I, se non ha una manifestazione fenotipica esterna) sopprime un gene ipostatico (solitamente indicato con B o b). L'epistasi recessiva si osserva quando l'allele recessivo di un gene epistatico inibisce l'espressione di uno qualsiasi degli alleli del gene ipostatico.

Anche la segregazione in base alle caratteristiche fenotipiche per ciascun tipo di queste interazioni è diversa. Con l'epistasi dominante, si osserva più spesso la seguente immagine: nella seconda generazione, la divisione per fenotipi sarà la seguente: 13:3, 7:6:3 o 12:3:1. Tutto dipende da quali geni si uniscono.
Nell'epistasi recessiva la divisione è la seguente: 9:3:4, 9:7, 13:3.
Complementarità
L'interazione di geni non allelici, in cui la combinazione di alleli dominanti di diversi tratti produce un nuovo fenotipo finora sconosciuto, è chiamata complementarità.
Ad esempio, questo tipo di reazione tra geni è più comune nelle piante (soprattutto nelle zucche).
Se il genotipo della pianta contiene un allele dominante A o B, la verdura riceve una forma sferica. Se il genotipo è recessivo, la forma del frutto è solitamente allungata.
Se nel genotipo sono presenti contemporaneamente due alleli dominanti (A e B), la zucca assume una forma a forma di disco. Se continui a incrociare (cioè continui questa interazione di geni non allelici con zucche di linea pura), nella seconda generazione puoi ottenere 9 individui con una forma discoidale, 6 con una forma sferica e una zucca allungata.
Tale incrocio consente di ottenere nuove forme ibride di piante con proprietà uniche.
Nell'uomo, questo tipo di interazione determina il normale sviluppo dell'udito (un gene è lo sviluppo della coclea, l'altro è lo sviluppo del nervo uditivo) e in presenza di un solo tratto dominante appare la sordità.
Polimerismo
Spesso la base per la manifestazione di un tratto non è la presenza di un allele dominante o recessivo di un gene, ma il loro numero. L'interazione di geni non allelici - la polimerizzazione - è un esempio di tale manifestazione.
L'azione polimerica dei geni può avvenire con o senza azione cumulativa. Durante il cumulo, il grado di manifestazione di un tratto dipende dall'interazione generale dei geni (più geni, più fortemente viene espresso il tratto). La prole con tale effetto è divisa come segue - 1: 4: 6: 4: 1 (il grado di espressione del tratto diminuisce, cioè in un individuo il tratto è espresso al massimo, in altri svanisce fino a scomparire completamente) .
Se non si osserva alcun effetto cumulativo, la manifestazione del tratto dipende dagli alleli dominanti. Se esiste almeno uno di questi alleli, si verificherà il tratto. Con un effetto simile, la scissione nella prole avviene in un rapporto di 15:1.
Azione dei geni modificatori
L'interazione di geni non allelici, controllati dall'azione dei modificatori, è osservata relativamente raramente. Un esempio di tale interazione è il seguente:

Tale interazione di geni non allelici negli esseri umani si verifica abbastanza raramente.
Pleiotropia
Con questo tipo di interazione, un gene regola l'espressione o influenza il grado di espressione di un altro gene.
Negli animali, la pleiotropia si manifestava come segue:
- Nei topi, un esempio di pleiotropia è il nanismo. Si è notato che incrociando topi fenotipicamente normali nella prima generazione, tutti i topi risultavano essere nani. Si è concluso che il nanismo è causato da un gene recessivo. Gli omozigoti recessivi smettevano di crescere e i loro organi interni e le loro ghiandole erano sottosviluppati. Questo gene del nanismo ha influenzato lo sviluppo della ghiandola pituitaria nei topi, il che ha portato ad una diminuzione della sintesi ormonale e ha causato tutte le conseguenze.
- Colorazione platino nelle volpi. La pleiotropia in questo caso si manifestava con un gene letale che, quando si formava un omozigote dominante, causava la morte degli embrioni.
- Negli esseri umani, l'interazione pleiotropica è dimostrata anche dall'esempio della fenilchetonuria
Il ruolo dell'interazione non allelica
In termini evolutivi, tutti i suddetti tipi di interazione dei geni non allelici svolgono un ruolo importante. Nuove combinazioni di geni causano la comparsa di nuove caratteristiche e proprietà degli organismi viventi. In alcuni casi questi segni contribuiscono alla sopravvivenza dell'organismo, in altri, al contrario, causano la morte di quegli individui che si distingueranno in modo significativo nella loro specie.

L'interazione non allelica dei geni è ampiamente utilizzata nella genetica dell'allevamento. Alcune specie di organismi viventi si preservano grazie a tale ricombinazione genetica. Altre specie acquisiscono proprietà molto apprezzate nel mondo moderno (ad esempio, lo sviluppo di una nuova razza di animali che ha maggiore resistenza e forza fisica rispetto ai suoi genitori).
Sono in corso lavori per utilizzare questi tipi di ereditarietà negli esseri umani al fine di eliminare i tratti negativi e creare un nuovo genotipo privo di difetti.
La manifestazione delle caratteristiche di un organismo dipende non solo dai geni ereditati, ma anche da come i geni interagiscono tra loro. Il genotipo contiene informazioni su determinati tratti, ma questi potrebbero non apparire nel fenotipo o apparire in modi diversi a seconda di come interagiscono i geni.
Interazione allelica
Ciascun cromosoma ha un cromosoma omologo ricevuto dall'altro genitore. I geni allelici che determinano tratti alternativi si trovano simmetricamente su questi cromosomi.

Riso. 1. Cromosomi omologhi.
Il modo in cui l'uno o l'altro tratto ereditario si manifesterà nel fenotipo dipende dal tipo di interazione genetica.
Dominazione
La dominanza può essere completa o incompleta.
In caso di dominanza completa appare il segno di un gene allelico chiamato dominante (A).
Un tratto alternativo è chiamato recessivo (a) e appare solo in assenza di uno dominante.
TOP 3 articoliche stanno leggendo insieme a questo
Con una dominanza incompleta appare un nuovo carattere intermedio. Ad esempio, in alcune piante il colore rosso (A) dei petali è dominante sul bianco (a).
Se, con la dominanza completa, i petali sono rossi (AA e Aa) o bianchi (aa), allora con l'eterozigote incompleto Aa avrà petali rosa.
Codominanza
Quando si eredita il gruppo sanguigno 4, funziona il principio della codominanza - quando i geni allelici Iᵇ e Iᵃ agiscono insieme e nessuno dei due è dominante o recessivo.
Sovraccarico
Se gli eterozigoti mostrano un tratto più forte di qualsiasi omozigote, allora questo tipo di interazione genetica è chiamata sovradominanza.
AA˂Aa˃aa
Ad esempio, la Drosophila ha geni che determinano la durata della vita. Il segno appare come segue:
- ahh- soggetti non vitali;
- aa- individui con aspettativa di vita normale;
- Ahh- aumento dell'aspettativa di vita.
Allelismo multiplo
In alcune popolazioni, i tratti sono codificati non da una coppia di geni allelici, ma da diversi alleli risultanti da mutazioni. Potrebbero esserci diverse dozzine di alleli di questo tipo.
In questo caso sono possibili diversi tipi di interazioni genetiche. I geni possono trovarsi in una relazione di dominanza completa o incompleta.
С ˃ сᵃ ˃ сᵇ ˃ с
Il gene C è dominante su qualsiasi gene, il gene cᵃ è dominante su tutti tranne C, ecc. Il gene c appare solo nello stato omozigote (cc).

Riso. 2. Allelismo multiplo nei conigli.
Interazione non allelica
Anche i geni non allelici si influenzano a vicenda.
Esempi di tali impatti sono:
- pleiotropia;
- epistasi;
- polimerismo;
- complementarietà.
L'effetto pleiotropico è l'influenza di un gene su diversi tratti. Ad esempio, i piselli dolci hanno lo stesso il gene determina:
- colore viola dei petali;
- pigmentazione della stipola;
- colore scuro del frutto.
L'effetto pleiotropico è molto diffuso in natura.
L'interazione epistatica è la soppressione dei geni di una coppia allelica da parte dei geni di un'altra coppia allelica.
Succede:
- dominante (A˃B);
- recessivo (aa˃B).
Con l'ereditarietà polimerica, diversi geni non allelici controllano un tratto e il grado della sua espressione può dipendere o meno dal numero di geni dominanti (effetto cumulativo).
L'interazione complementare è anche chiamata aggiuntiva, perché con essa i geni non allelici determinano congiuntamente il tratto. Ciò può accadere anche se uno o entrambi individualmente non codificano tale funzionalità.